|
新闻动态
|
科研进展
东亚喀斯特常绿阔叶林发展历史对来江藤属植物演化及扩散的影响 文章来源:中国科学院东亚植物多样性与生物地理学重点实验室 | 发布时间:2023-05-30 | 作者:陈哲等 | 浏览次数: | 【打印】 【关闭】 东亚植物区系物种丰富且具有大量特有种,其演化受到新生代(~6500万年)以来的地质、气候事件的深刻影响。该地区是北半球物种演化的重要中心,也是科学研究的热点地区之一。来江藤属(Brandisia)以中国西南喀斯特地区为分布中心,个别物种可分布到东南亚,如缅甸、泰国、越南等(图1)。该属为列当科(Orobanchaceae)成员,具有(一定的)半寄生性。植株一般为半灌木,具有一定攀缘性,因而也多少表现出藤本特性(图2)。该属植物主要生长于中低海拔(500~3000 m)的山地林下、林缘、山坡、路边、河岸等较开阔(具有一定干扰)的生境中,是东亚喀斯特常绿阔叶林中的典型伴生类群。 中国科学院昆明植物研究所高山植物多样性研究团队通过广泛取样构建了迄今最为完整的来江藤属的系统发育关系,结合分化时间估算、祖先分布区推断、东亚常绿阔叶林发展历史以及来江藤属植物的生物学特性,深入探讨了该属的起源、演化及扩张等科学问题,丰富了对东亚植物区系的理解。
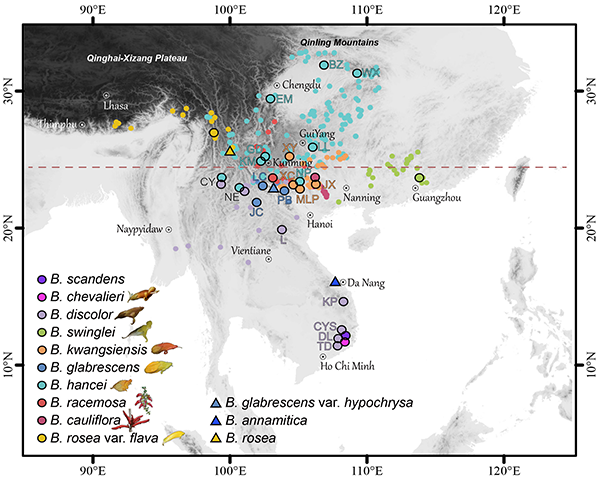
图1. 来江藤属各物种的地理分布及本研究的采样点。具有黑色轮廓的点为采样点。三角形表示有少量标本记录,但未能收集到的物种。红色虚线表示北回归线(23.5 N°)。
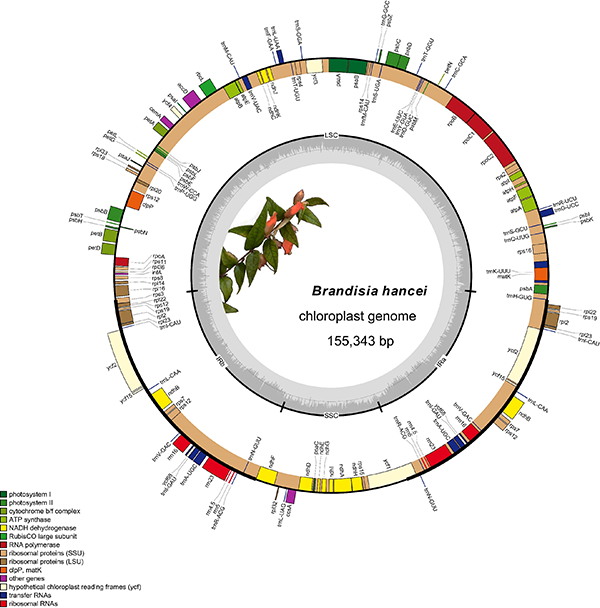
图2. 来江藤属代表植物。A. 红花来江藤黄花变种(Brandisia rosea var. flava C.E.C. Fisch.);B. 茎花来江藤(B. cauliflora P.C. Tsoong & L.T. Lu);C. 总花来江藤(B. racemosa Hemsl.);D. 来江藤(B. hancei Hook. f.);E. 退毛来江藤(B. glabrescens Rehder);F. 广西来江藤(B. kwangsiensis H.L. Li);G. 岭南来江藤(B. swinglei Merr.);H. 异色来江藤(B. discolor Hook. f. & Thomson)。 研究结果显示,虽然来江藤属植物具有半寄生特性,但其叶绿体基因组结构保守,基因数目完整、无假基因(图3),这与列当科中的大多数寄生、半寄生类群不同(其叶绿体基因通常发生严重重排、简化等)。这可能是由其生活型决定的,来江藤属植物为列当科中少有的(亚)灌木,体型较大(调查发现异色来江藤可高达3米,呈较大藤本状),这可能要求其有较高的光合效率,因而强烈的自然选择维持了其叶绿体基因组的保守性。
图3. 来江藤(Brandisia hancei)叶绿体全基因组图谱。 分子系统学研究结果显示,来江藤属为单系类群。由于存在核质冲突,该属在列当科中的位置不稳定(图4)。属下种间关系大体一致,形成与形态吻合的三个支系:1)花萼为二唇形的红花来江藤;2)呈总状花序的茎花来江藤和总花来江藤;3)其他物种。越南特有的两个物种(B. chevalieri Bonati和B. scandens Bonati)嵌在异色来江藤内部;且其本身的单系性没有得到很好的支持。此外,在叶绿体系统发育树中,异色来江藤不能形成单系,来自越南和老挝的样本形成一支,但与中国分支分开。进一步对标本和文献进行仔细查阅后,研究人员认为B. chevalieri和B. scandens不成立,应为异色来江藤的异名。
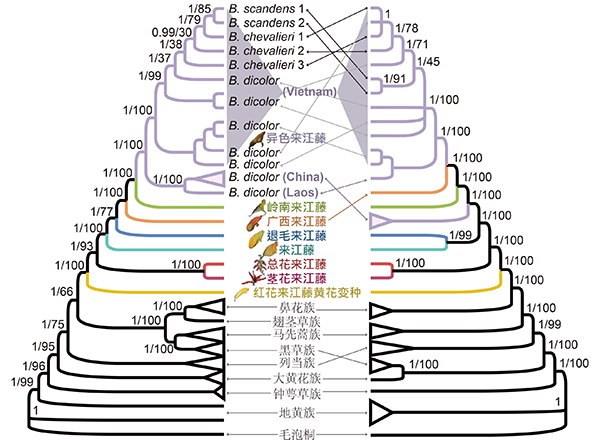
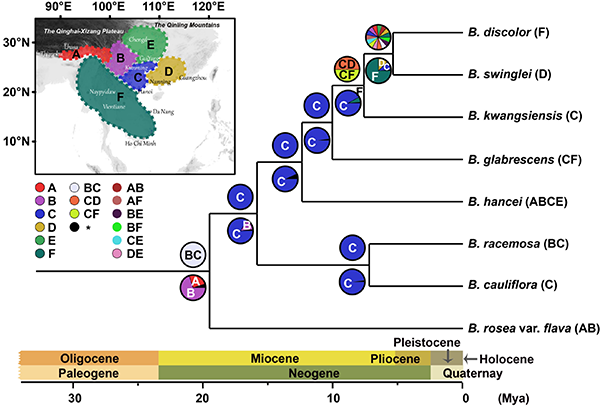
图4. 来江藤属系统位置及属下各物种的系统关系。左图为基于四个核基因片段(3S/ITS, PHYA, PHYB_1 and PHYB_2)的系统框架;右图为基于叶绿体基因组的系统框架。节点处数值分别表示贝叶斯后验概率和Bootstrap支持率。 分化时间估算及祖先起源地推断显示来江藤属可能起源于早渐新世(约3300万年前),起源地可能为东喜马拉雅至中国西南一带。该属物种可能于早中新世(约1900万年前)在喀斯特常绿阔叶林中开始分化(图5和6)。基于来江藤属的分布格局、分化时间、其半寄生和藤本特性以及东亚植物区系的发展历史等信息,认为喀斯特常绿阔叶林的起源与发展(可能在渐新世-中新世之交(约2300万年前)或更早)是影响该属物种分化、扩张的重要因素,为来江藤属植物提供了可能的植物寄主与合适的生境,从而促进了该属的分化,使得其在中国西南部及越南北部形成现代分布中心。
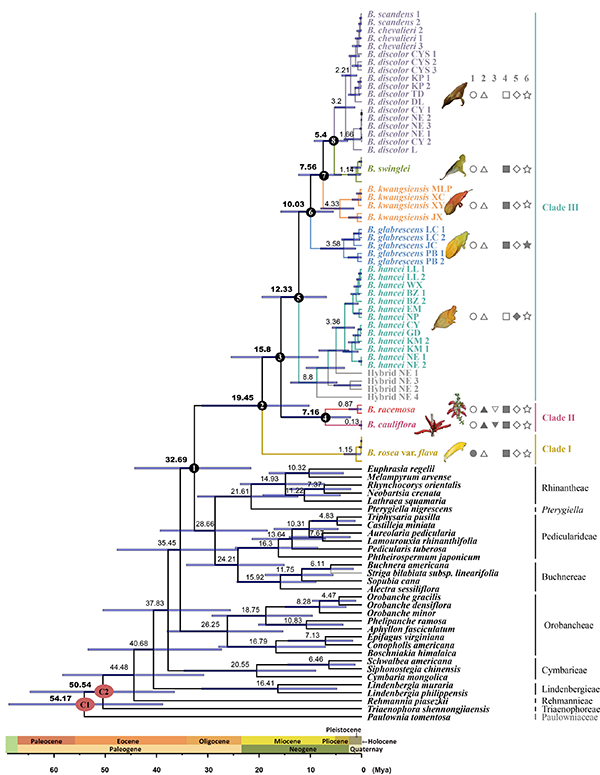
图5. 来江藤属及相关类群分化时间推断(BEAST)。采用二次标定方法,两个红色节点为标定点。特征1,●花萼二唇形,○花萼五裂;特征2,▲总状花序,△单花;特征3,▼紧凑花序生于主茎上,▽总状花序生于枝端;特征4,■花季夏(秋),□花季冬(春);特征5,◆叶基心形,边缘向后反卷,◇叶基楔形、近圆形,边缘不反卷;特征6,★花萼大、亮黄色,☆花萼小、灰色或棕色。
图6. 来江藤属祖先区域重建。采用S-DIVA(枝上)和BBM(枝下)法。A,东喜马拉雅;B,云南高原;C,中国西南喀斯特核心区;D,华南;E,华中; F,中南半岛。 该属起源较早,但分化较晚,现存物种多样性低,且分布范围有限,可能的原因有:1)其姐妹群可能已灭绝,导致对该属的起源时间估算过早;2)该属可能为孑遗类群,目前的狭窄分布可能是其分布区受历史上气候变化的影响而收缩的结果;3)该属主要分布在中国西南和越南北部,此地区自第三纪以来环境较为稳定、适宜,这可能导致了较低的物种形成速率和扩散速率;4)该属为灌木,而木本植物因其寿命和世代周期较长而表现出较低的多样化速率;5)该属物种均为鸟媒植物,传粉鸟类的分布(主要在温暖的热带、亚热带地区)也是影响来江藤属植物分布的重要因素。 该研究以Historical development of karst evergreen broadleaved forests in East Asia has shaped the evolution of a hemiparasitic genus Brandisia (Orobanchaceae)为题在线发表于Plant Diversity。中国科学院昆明植物研究所陈哲博士为论文第一作者,孙航研究员和牛洋研究员为共同通讯作者。越南科学技术院的合作者参与了该研究。 (责任编辑:李雪)
|

版权所有 Copyright © 2002-2025 中国科学院昆明植物研究所,All Rights Reserved 【滇ICP备05000394号】
地址:中国云南省昆明市蓝黑路132号 邮政编码:650201
点击这里联系我们